 Suche
Suche
 Mein Konto
Mein Konto
Bakteriene vi puster inn hver dag

I en fersk studie publisert i tidsskriftet PNAS, undersøkte forskere globale luftbårne bakteriesamfunn for å forstå deres samfunnsstruktur og biogeografiske distribusjonsmønstre. I tillegg undersøkte de deres interaksjoner med andre jordmikrobiomer, spesielt overflatehabitater. Læring: Globalt luftbårent bakteriesamfunn - Interaksjoner med jordens mikrobiomer og menneskeskapte aktiviteter. Bildekreditt: Lightspring/Shutterstock Bakgrunn Atmosfæren er det mest uberørte mikrobielle habitatet på jorden, og luftbårne bakterier er de mest komplekse og dynamiske samfunnene som påvirker jordens mikrobiomer. Det er mer enn 1 × 104 bakterieceller/m3 og hundrevis av unike taxa i luften. Storskala studier har...

Bakteriene vi puster inn hver dag
I en studie nylig publisert i tidsskriftet PNAS Forskere undersøkte globale luftbårne bakteriesamfunn for å forstå deres samfunnsstruktur og biogeografiske distribusjonsmønstre. I tillegg undersøkte de deres interaksjoner med andre jordmikrobiomer, spesielt overflatehabitater.

bakgrunn
Atmosfæren er det mest uberørte mikrobielle habitatet på jorden, og luftbårne bakterier er de mest komplekse og dynamiske samfunnene som påvirker jordens mikrobiomer. Det er mer enn 1 × 104 bakterieceller/m3 og hundrevis av unike taxa i luften. Storskala studier har systematisk dokumentert de mikrobielle egenskapene i jordsmonn, hav og menneskelig avfall. De har også foreslått en sammenheng mellom luftbårne mikrobiomer og overflatemiljøer. Imidlertid er det mangel på studier som dokumenterer luftbårne mikroorganismer, spesielt angående deres samfunnsstruktur.
Mikrober lever ikke isolert. I stedet har de flere økologiske forhold som spenner fra gjensidighet til konkurranse. Derfor kan å bestemme deres biogeografiske distribusjonsmønstre og interaksjoner med andre jordmikrobiomer som definerer deres opprinnelse kaste lys over effektene av klima-/miljøendringer og menneskeskapte aktiviteter.
Om å studere
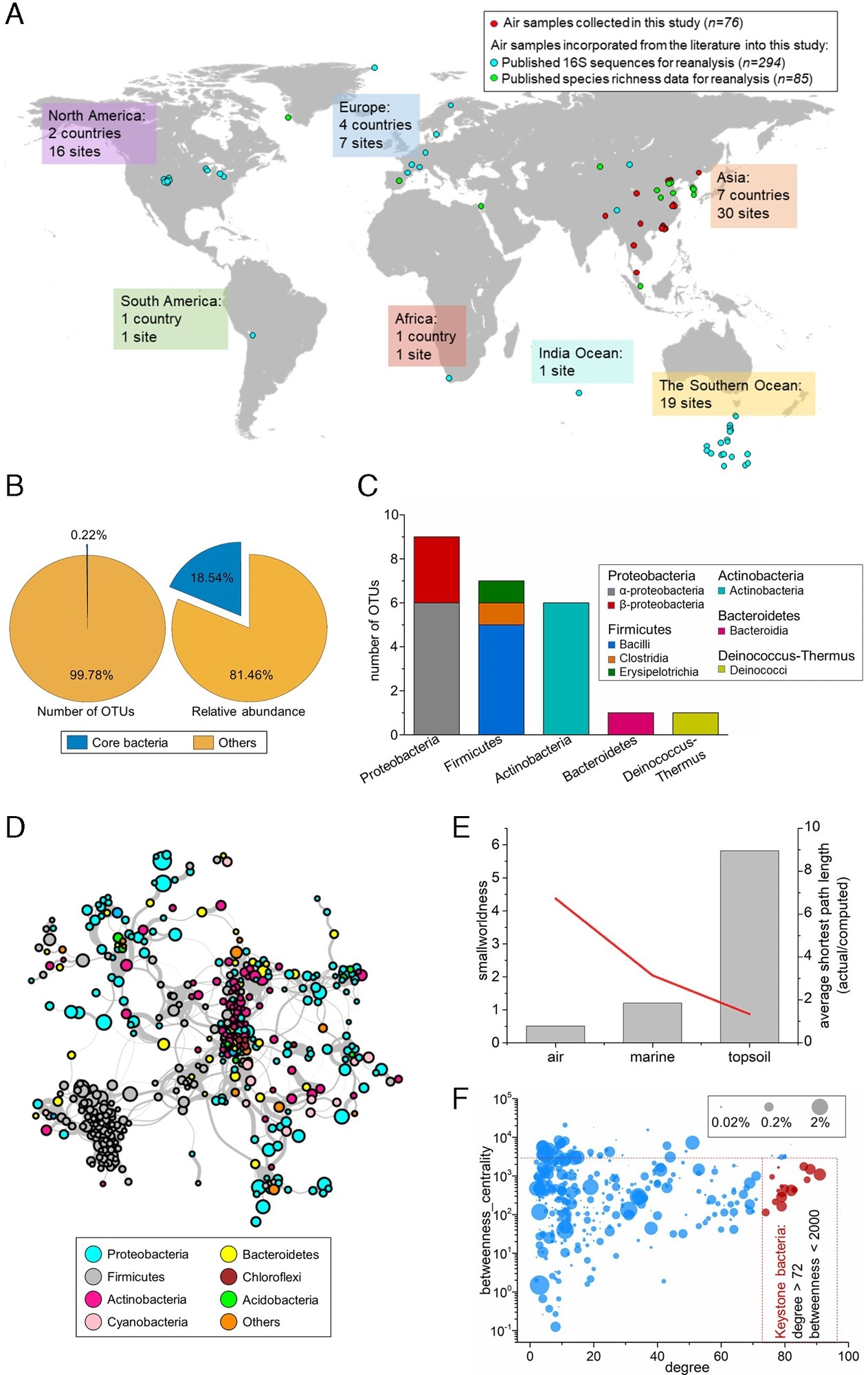
I denne studien utviklet forskerne først et globalt datasett om luftbårne bakterier for å vurdere graden av fellesskap og sammenheng. Dette datasettet inkluderte 76 nylig innsamlede luftpartikkelprøver kombinert med 294 prøver samlet inn for tidligere studier på 63 steder over hele verden. Prøvetakingssteder varierte i høyde og geografi og inkluderte bakkenivå, hustak (fra 1,5 m til 25 m i høyden) opp til fjell 5 380 m over havet, tett befolkede urbane byer og den avsidesliggende polarsirkelen.
Teamet innhentet datasettet for sammenligning fra Earth Microbiome Project (EMP), som har samlet inn mer enn 5000 prøver fra 23 overflatemiljøer. Den luftbårne bakteriereferansekatalogen inneholdt mer enn 27 millioner ikke-redundante 16S ribosomale RNA (rRNA) gensekvenser.
I tillegg konstruerte forskere et globalt luftbårent fellesskapsnettverk som inkluderte 5 038 signifikante korrelasjonsforhold (Spearmans ρ > 0,6) mellom 482 assosierte operasjonelle taksonomiske enheter (OTUs). OTU-er er analytiske enheter gruppert etter DNA-sekvenslikhet i mikrobiell økologi. Til slutt brukte teamet strukturell ligningsmodellering (SEM) for å undersøke mekanismene som driver mikrobielle samfunn. De beregnet også den samlede effekten av miljøfiltre og bakterielle interaksjoner på samfunnsdesign.
Studieresultater
10 897 taxa ble påvist fra 370 individuelle luftprøver, og de fleste bakteriesekvenser tilhørte fem phyla. Firmicutes, Alphaproteobacteria, Gammaproteobacteria, Actinobacteria og Bacteroidetes utgjorde henholdsvis 24,8 %, 19,7 %, 18,4 %, 18,1 % og 8,6 % av disse bakteriesekvensene. Abundance-occupancy-forholdet (AOR) mellom prøvene okkupert av en bakteriell takson og dens gjennomsnittlige masse i den globale luften viste en sigmoid-kurve, lik det observerte mønsteret for fordelingen av dyreliv og planter på jorden.
Luft er et frittflytende, dynamisk økosystem som muliggjør langdistansetransport av bakteriesamfunnene den støtter. Imidlertid så det ut til at bakteriesamfunnet var godt knyttet til lokalmiljøet, spesielt kildebidragene og luftkvalitetsforholdene som følge av menneskeskapte aktiviteter. Reduserte filtreringseffekter fra miljøet og økte menneskerelaterte kildebidrag har ført til lavere biomassebelastninger, høyere frekvenser av patogene bakterier og mer destabiliserte nettverksstrukturer.
Spesielt var luftbårne bakterier ikke tett forbundet sammenlignet med deres motparter i matjord og marine miljøer og hadde en gjennomsnittlig intranode-tilkobling på 5,24. De hadde en tilfeldig klyngingstilnærming, og topologien hadde lav motstand mot endringer. De observerte fjerne relasjonene og løse klynger i nettverket antydet at det er mer sannsynlig at det luftbårne bakteriesamfunnet blir forstyrret avhengig av miljøforholdene, som vanligvis fører til drastiske endringer i bakteriesammensetningen. Funksjonene til atmosfæriske bakterielle taxaer har blitt utledet basert på deres genetiske informasjon i andre habitater.
Omics e-bok
Sammenstilling av de beste intervjuene, artikler og nyheter fra det siste året. Last ned en gratis kopi
Teamet fant potensielle assosiasjoner mellom luftbårne bakteriesamfunn og andre mikrobielle habitater på overflaten. Den estimerte totale mengden av luftbårne bakterier (1,72 × 1024 celler) var sammenlignbar med hydrosfæren og en til tre størrelsesordener lavere enn i andre habitater (f.eks. jord).
Av de 23 store jordhabitatene som ble undersøkt i den nåværende studien, viste terrestrisk luft større likhet med menneske- og dyremiljøer, mens offshoreluft viste et nærmere forhold til havsystemer. Videre viste evalueringer basert på Bayesianske metoder at egenskapene til det tilsvarende overflatemiljøet bestemte de dominerende kildene til luftbårne bakterier. Spesielt bidro menneskelige kilder mer til luftbårne bakterier i urbane områder, spesielt på landbaserte steder, et funn som stort sett ble ignorert i tidligere utslippsmodelleringsstudier.

Forfatterne fant ingen signifikante forskjeller i rikdommen av luftbårne bakteriesamfunn mellom urbane og naturlige områder innenfor samme breddegradsområde. Geografisk plassering spilte imidlertid en rolle. Ensartetheten til bakteriesamfunnene i byluften var betydelig lavere. For eksempel var den relative forekomsten av patogene arter, Burkholderia og Pseudomonas, høyere i urbane områder enn i naturområder (5,56 og 2,50 % mot 1,44 og 1,11 %). I tillegg bidro bakterier mindre til partikkelmasse (PM) i urbane områder enn i naturområder, noe som tyder på at urbanisering økte andelen ikke-biologiske partikler i luften (f.eks. støv).
Patogenene med høyest dødelighetsrisiko, Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa og Enterobacter arter (ESKAPE), var mer vanlige i byluft. Samforekomstnettverket av urbane luftbårne bakteriesamfunn antydet at menneskeskapte påvirkninger destabiliserte nettverksstrukturen deres, noe som igjen også endret den bakterielle taksonomiske sammensetningen.
Forfatterne fant at flere faktorer påvirket luftbårne bakteriesamfunn – for eksempel geografiske steder sammen med typiske miljøfaktorer. De biotiske interaksjonene mellom keystone og kjernebakteriesamfunn og bakteriell rikdom interagerte betydelig. Av alle deterministiske prosessene var miljøfiltrering den viktigste determinanten for strukturen og fordelingen av luftbårne mikrobielle samfunn.
Konklusjoner
Oppsummert stammer nesten 46,3 % av luftbårne bakterier fra miljøet, og stokastiske prosesser formet først og fremst fellesskapsdannelse. Videre har det kjennetegn ved luftbårne bakterier i urbane områder vært den økende andelen potensielle patogener fra menneskerelaterte kilder. Til slutt påvirket luftbårne bakteriekildeprofiler en betydelig høyere prosentandel av strukturelle variasjoner enn for luftkvalitet og lokale meteorologiske forhold (43,7 % versus 29,4 % og 25,8 %), vurdert ved variasjonsfordelingsanalyse (VPA).
Referanse:
- Globale luftgetragene Bakteriengemeinschaft – Wechselwirkungen mit den Mikrobiomen der Erde und anthropogenen Aktivitäten, Jue Zhao, Ling Jin, Dong Wu, Jia-wen Xie, Jun Li, Xue-wu Fu, Zhi-yuan Cong, Ping-qing Fu, Yang Zhang, Xiao- San Luo, Xin-bin Feng, Gan Zhang, James M. Tiedje, Xiang-dong Li, PNAS 2022, DOI: https://doi.org/10.1073/pnas.2204465119, https://www.pnas.org/doi/full/10.1073/pnas.2204465119
.